
热门关键词: ELISA试剂盒 - 大鼠ELISA试剂盒- 小鼠ELISA试剂盒
全国统一客服热线:
0755-28715175
热门关键词: ELISA试剂盒 - 大鼠ELISA试剂盒- 小鼠ELISA试剂盒
全国统一客服热线:
0755-28715175


T 细胞通过其抗原受体的激活与它们所识别的同源抗原的识别强度密切相关,这种强度由可变互补决定区环所决定。
2026年2月13日,芝加哥大学Erin J. Adams团队在Science Immunology 在线发表题为TCRγ constant usage tunes human γδ T cell antigen sensitivity, thymic programming, and peripheral function的研究论文。
该研究发现,在人类γδ T 细胞受体(TCRs)中,γ恒定结构域独立于可变区域抗原识别来调节激活强度。通过单细胞 RNA 测序数据,该研究证明了体内 Cγ 的使用与不同的表型相关,使用 Cγ1 的细胞在胸腺中具有更成熟的细胞毒性效应表型,并在周围组织中具有更高的颗粒酶表达。
具有 Cγ1 的 TCR 也在结直肠肿瘤中被选择性扩增。Cγ2 的使用与发育中的初始表型相关,与周围组织中的抑制和伤口愈合相关。总之,该研究提出,使用 Cγ1 或 Cγ2 对激活的调节会导致γδ T 细胞表型和克隆扩增在其整个生命周期中的差异,为人类γδ T 细胞提供了另一个“调节旋钮”来调节其功能。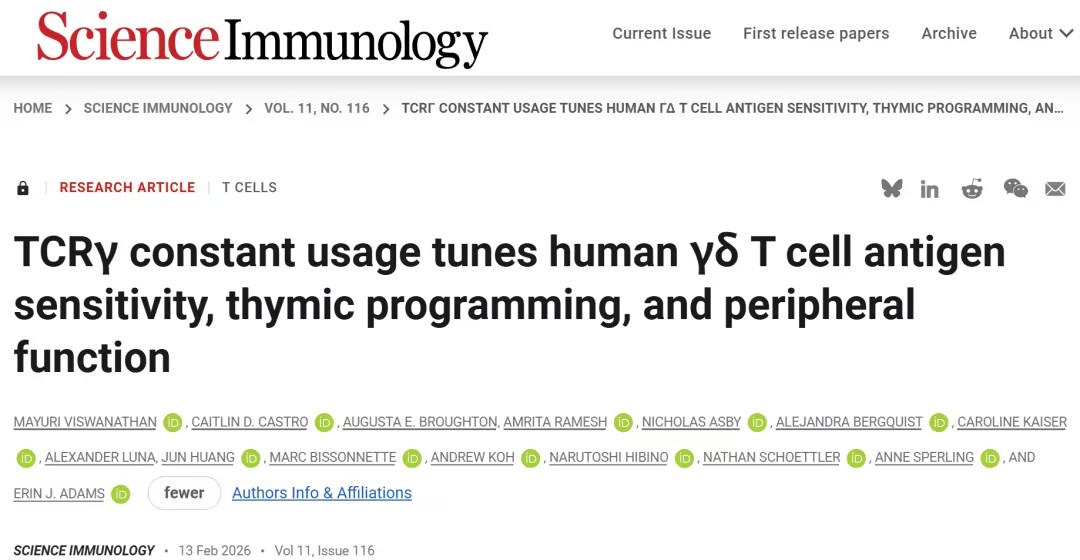
Gamma delta(γδ)T 细胞在先天免疫和适应性免疫的交界处发挥着作用。尽管它们在人类外周血中的比例较小,但在屏障组织中却可能成为 T 细胞的主体。γδ T 细胞在一系列免疫和非免疫功能中发挥着关键作用,从抗病毒反应、抗肿瘤反应到伤口愈合和记忆等。人类的 γδ T 细胞在几个关键方面与小鼠的 γδ T 细胞不同,包括可变基因的使用、抗原识别和功能极化,这使得了解这些细胞在人类中的特征变得颇具挑战性。
αβ 和 γδ T 细胞受体(TCRαβ 和 TCRγδ)均由可变域和恒定域组成。可变域包含互补决定区(CDR)环,而恒定域则包括连接肽(CP)区域和跨膜区域。在 TCRαβ 中,α 和 β 链通过位于靠近细胞膜处的两个保守半胱氨酸残基之间的二硫键进行共价连接。
为了在细胞表面稳定表达并进行细胞内信号转导,这种异二聚体必须与 CD3 蛋白 ε、γ、δ 和 ζ 相结合。尽管 TCR 和 CD3 蛋白的跨膜区域在该 TCR-CD3 复合物的组装中起着重要作用,但 TCRαβ 恒定域在信号转导中也至关重要,因为恒定域 CP 会与 CD3 成分结合并稳定膜邻近的相互作用。
有趣的是,β链和γ链的恒定结构域都可以由两个独立的基因来编码。β链恒定基因 TRBC1 和 TRBC2 分别编码 Cβ1 和 Cβ2,它们在细胞外部分仅相差四个氨基酸,从结构和功能上看是可以相互替代的。Cβ2 在外周血中健康αβ T 细胞的 TCR 中的使用比例更高,但 Cβ1 和 Cβ2 都存在于 CD4 和 CD8 的每个功能亚群的 TCR 中,这表明 TCR 恒定β链在 TCR 中的使用是在胸腺发育过程中随机选择的,并且不会影响 T 细胞的下游功能。
然而,与β链不同的是,TCRγδ的γ链恒定基因编码的蛋白质在它们所推测的细胞外结构上有所不同,最显著的是 Cγ1 和 Cγ2 的 CP 区域长度存在差异(分别由 TRGC1 和 TRGC2 基因编码)。
γδ T 细胞中 Cγ1 和 Cγ2 的使用模式及功能模型(图源自Science Immunology )
最近的结构研究揭示了含 Cγ1 和含 Cγ2 的 TCR 之间的激活差异。具有相同抗原特异性的 TCR,在经过 Cγ1 修饰后比经过 Cγ2 外显子 2 重复等位基因修饰的 TCR 能获得更高的激活程度。
另外一些研究还探讨了αβ 和γδ T 细胞之间 CP 的差异,结果表明γδ CP 的长度增加可能为受体提供了更多的灵活性,并降低了 TCRγδ 配体的敏感性。然而,仍有疑问在于这些发现是否适用于 TRGC 基因的外显子 2 三重等位基因,以及这些激活差异是否会导致持久的功能性影响。
这些数据表明,TRGC 的使用可能在结构上影响γδ T 细胞的抗原反应和功能极化。研究人员首先在一个简化模型中,使用已知抗原特异性的重组 TCR 进行测试,然后在人类组织中使用基于聚合酶链反应(PCR)的方法进行测试,最后在生成的和先前发表的单细胞 RNA 测序(scRNA-seq)数据集中进行测试。研究人员发现,TRGC 的使用调节了激活过程,并与功能表型和克隆扩增的偏差相关,在健康和疾病状态下均如此。
0755-28715175/33164177

粤公网安备 44030902000304号

